

While growing up, I had a friend named Tom who had an unusual older brother named Tim. Back then, Tom and I really looked up to Tim. Maybe that’s why whenever I think of my friend Tom, I can’t help but also think about Tim. Tom and Tim have become inseparable in my mind.
TOM and TIM are also inseparable when it comes to the biogenesis of mitochondria. In this process, TOM and TIM aren’t brothers, but large protein complexes found in the outer and inner membranes of mitochondria. TOM (translocase of the outer membrane) and TIM (translocase of the inner membrane) function as biomolecular machines that operate in tandem to import newly made proteins into mitochondria.
The inseparability of TOM and TIM represents a significant challenge to the endosymbiont hypothesis—a cornerstone idea in evolutionary biology that offers an explanation for the origin of organelles, such as mitochondria.
How Complex Is Protein Transport into Mitochondria?
The similarity between organelles and bacteria serves as the chief line of evidence for the endosymbiont hypothesis. For example, mitochondria—thought to have descended from α-proteobacteria—are about the same size and shape as a typical bacterium and have a double membrane structure similar to gram-negative microbes. These organelles also divide in a way that is reminiscent of bacterial cells.
From my vantage point, similarities between mitochondria and α-proteobacteria are not enough to establish the validity of the endosymbiont hypothesis. Evolutionary biologists must also explain how mitochondria became fully integrated into the host cell’s metabolic systems. Even though biochemists are figuring out how the process of protein transport works, they must also identify a plausible evolutionary pathway that can adequately account for the evolution of this biochemical operation.
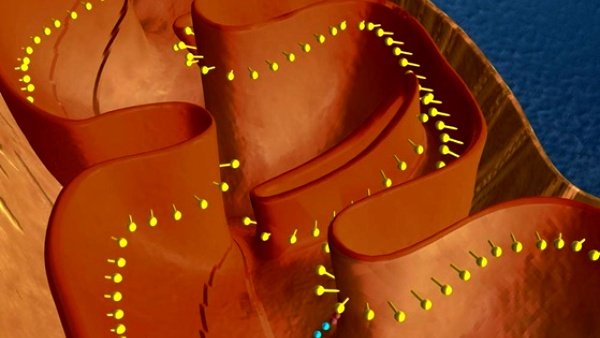
Except for select proteins, most mitochondrial proteins are made in the cytosol of the cell and transported into the mitochondria. The overall process of mitochondrial protein biogenesis consists of four stages: (1) protein synthesis (2) targeting the protein to the mitochondria; (3) transporting the protein into the mitochondrial lumen; and (4) targeting the protein to its final destination in the organelle.
The cell’s machinery initially makes mitochondrial proteins as pre-proteins with a signal sequence at one of its ends (the N terminus). The signal sequence has a specialized structure (an amphipathic α-helix) that serves to target the proteins to mitochondria. Think of the signal sequence as analogous to an address label that tells the post office where to deliver a letter. Receptor proteins that are part of the TOM complex recognize the signal sequence and transport the protein through a channel within the TOM interior into the intermembrane space (the region between the mitochondrian’s inner and outer membranes). Proteins dubbed chaperones keep the mitochondrial proteins unfolded and stabilized throughout this process.
Once in the intermembrane space (the region between the outer and inner membranes), two different TIM complexes (TIM22 and TIM23) work together, taking the protein “baton” from the TOM complex and ushering the protein into the lumen (or the matrix) of the mitochondria. If the protein is to remain within the lumen (because that’s where it performs its work), then proteins called peptidases remove the signal sequence and the protein adopts its intended three-dimensional shape.
If the protein is to be incorporated into the inner membrane, it possesses an additional targeting sequence that is recognized by another protein complex dubbed OXA. This biomolecular ensemble inserts the protein into the inner membrane.
If the protein is to carry out its work in the intermembrane space, then the OXA complex will transport the protein back across the inner membrane. Alternatively, some proteins destined to operate in the inner membrane space possess a stop signal sequence. These sequences prevent the TIM22 and TIM23 complexes from transporting it across the inner membrane into the lumen. Instead, peptidases in the intermembrane space remove the signal sequence, allowing the protein to adopt its operational structure.
Finally, if the protein is to be incorporated into the outer membrane, then another complex referred to as SAM inserts it into the outer membrane.
The Challenge to the Endosymbiont Hypothesis
Each stage of mitochondrial protein biogenesis involves multiple steps with each one carried out by an ensemble of proteins. Moreover, each step of the process must be precisely integrated with the other steps. If not, the entire process of mitochondrial protein biogenesis fails. To put it another way, each step of the process involves an irreducibly complex biochemical apparatus which, in turn, integrate with each other to form the irreducibly complex process of mitochondrial protein biogenesis. That is, mitochondrial protein biogenesis can be characterized as an integrated, hierarchal, multilayered ensemble of irreducibly complex systems.
For the mitochondrial protein biogenesis to emerge from an evolutionary standpoint, a number of biochemical systems had to simultaneously evolve and become integrated with one another. For example, once mitochondrial genes became incorporated into the host genome, DNA sequences specifying signal sequences had to evolve and become precisely appended to every one of the mitochondrial DNA sequences. The TOM, TIM22, and TIM23 complexes had to evolve simultaneously to recognize mitochondrial proteins and work in tandem to move proteins into the mitochondria. In addition, chaperones had to emerge that would recognize mitochondrial proteins and keep them unfolded during the transport process. Signal peptidases had to evolve to remove signal sequences from mitochondrial proteins with exacting precision. Finally, stop sequences and additional targeting sequences had to evolve and become precisely positioned within the mitochondrial protein genes.
In effect, no one knows how mitochondrial protein biogenesis could have evolved. According to cell biologist Franklin Harold, “The origin of the machinery for protein import is more complicated and is subject to much debate.”1 Harold also states, “Most of the transferred genes continue to support mitochondrial functions, having somehow acquired the targeting sequences that allow their protein products to be recognized by TOM and TIM and imported into the organelle.”2 To say that “the origin of the machinery for protein import” is a “complicated” system that “somehow” evolved is not a scientific explanation for how this complex biochemical system arose. In the absence of a plausible evolutionary route for mitochondrial protein transport, it is reasonable to be skeptical of the endosymbiont hypothesis.
A Creation Model Perspective on the Origin of Mitochondria
While evolutionary biologists view the similarities between mitochondria and α-proteobacteria as evidence for the endosymbiont hypothesis, it is possible to view these similarities from a creation model vantage point as shared design features based on an archetypal design.
As I pointed out earlier, mitochondrial genomes exhibit an exquisite biochemical logic that undergirds their structure and function, making it all the more reasonable to view these organelles as the Creator’s handiwork.3 Bolstering this conclusion is the multi-tiered irreducible complexity of protein mitochondrial biogenesis. As I discussed in The Cell’s Design, irreducible complexity is a hallmark feature of many human designs and should be viewed as an indicator of intelligent design.